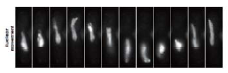
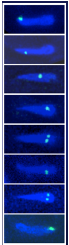
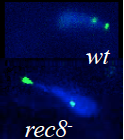
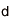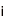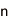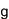ヒトを含むすべての真核生物は、生殖細胞(精子や卵子等)を作り出す際に、減数分裂と呼ばれる特殊な細胞分裂を行っています。この減数分裂の際に、遺伝情報を担う染色体が核の特定の場所に集合し、父母に由来する同種の2 本の染色体(相同染色体)が接合(対合)し、二価染色体を形成することで、遺伝情報の組換えを効率的に行います。これは、生命に進化と多様性をもたらす生物学的に極めて重要な現象です。
一方、相同染色体の対合が正常に行われないと、染色体異常に起因する病気(ダウン症等)や流産の原因にもなります。このため、対合のメカニズムは、医学的見地からも解明が長い間求められてきました。しかし、どのようにして相同染色体が壊れやすい遺伝情報を守りつつ、対合する相手を認識し、組換え部分で接合するかということについて、これまで明らかにされていませんでした。
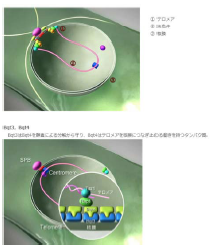
")
細胞質融合")
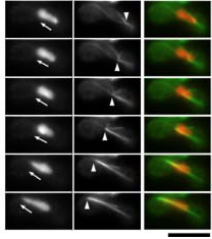
テロメアが核膜上の一か所に集合")
核融合")
核の往復運動による相同染色体の近接と接着")
sme2を起点に相同染色体が対合")

")
 染色体数1 セット2 本のみの簡易モデル")