| |  |
| | |
 | ||||
| 大岩 和弘 | ||||
| 生き物は動く 大型の動物からプランクトンのような微小な生物まで、生き物には“動き”があります。筋肉や鞭毛・繊毛は生物の動きを作り出すための装置です。この装置の中を詳細に見ていくと、力を発生する本体は「タンパク質モータ」と呼ばれるタンパク質であり、これらが無数に集まって“動き”を作り出している事がわかってきました。 タンパク質モータは、生物共通のエネルギー通貨であるアデノシン3リン酸(ATP)を加水分解し、そこから得られるエネルギーを使ってタンパク質繊維との間で滑り運動を行ないます。10ナノメートルほどの大きさのタンパク質モータは、タンパク質繊維の上をリニアモータのように滑走するのです。 | ||||
鞭毛運動のモータ・ダイニン
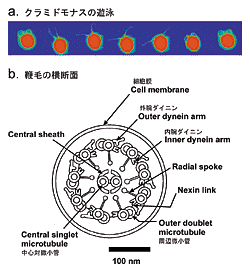
鞭毛の横断面を見ると、割り箸を束ねたような構造が見られます(図1.b)。中央に2本の微小管(中心対微小管)があり、9本の微小管(周辺微小管)がそれらを取り囲むように並んで、鞭毛全長にわたって伸びています。この構造は“9+2構造”と呼ばれ、ほとんどの生物の鞭毛・繊毛に共通してみられるものです。周辺微小管から隣の微小管に突き出しているのがダイニン腕と呼ばれる構造で、内腕と外腕の2列が微小管の上に規則的に配列しています。このダイニン腕はタンパク質モータとダイニンが複数集まってできた複合体で、隣の周辺微小管との間で力を出し“滑り”を生じて鞭毛の屈曲を作り出すのです。ダイニンはタンパク質モータの中で最も大きく、その存在は早くから知られていました。しかし、その分子構造の複雑さから解明されていない点が数多く残っています。 | ||||
ダイニン1分子を測る
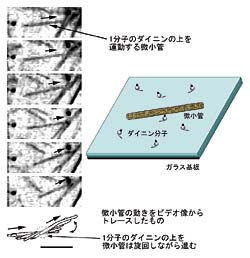
次にダイニンをガラス基板上に低密度で吸着させて機能測定を行ないます。微小管を加えると、ダイニンと結合した微小管は滑り運動をはじめます。運動している微小管を観察すると、ガラス基板上の1点で支えられ左右に旋回しながら運動しているものが見つかります。この旋回運動は熱揺動によるものです。支点にはまさにダイニン1分子がいて微小管の連続的な運動を起こしているのです(図3)。ダイニン分子に結合してから離れるまで微小管が滑った距離は平均で3.6マイクロメートルでした。
筋肉の収縮を引き起こすミオシンや、神経細胞内の物質輸送を行うキネシンなどのタンパク質モータ分子は、2つのモータ領域を持っています。タンパク質モータは、これらのモータ領域をタンパク質繊維に交互につけたり離したりして、二足歩行のように進むと考えられています。連続的な運動のためには、タンパク質モータはタンパク質繊維に複数の結合部位で結合してそこから離れないようにすることが必要であると考えられるのです。 ミオシンやキネシンはたいへん良く似た構造を持つことから、このような運動形式はタンパク質モータに普遍的なものとされ、運動機構が明らかになっていなかったダイニンにもあてはまるものと考えられていました。 すなわち、2つ以上のモータ部分を持たないタンパク質モータは、1分子では長い距離を動くことはできないとされていたのです。しかし、このダイニンはモータ領域を1つしか持たないのに、タンパク質繊維から離れることなく自分自身の大きさの300倍を超える距離を1分子で運動したのです。 この発見は、これまでに提唱されている生物運動のメカニズムの定説ではまったく説明できないものです。 | ||||
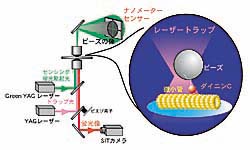
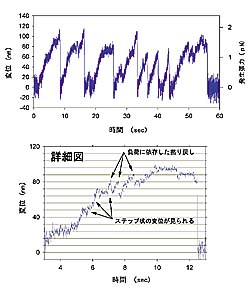
| レーザートラップ タンパク質モータを捕捉しその挙動を分子1個で直接計測する試みとして、レーザートラップを用いた力学計測があります。レーザートラップは対物レンズで集光したレーザーによって、微小粒子を捕捉・操作するものです。このレーザートラップに捕捉した微小粒子の小さな位置変化を測定すれば、微小粒子にかかる力を測ることができます。 ダイニン1分子を吸着させた微小粒子をレーザートラップで捕捉し微小管と接触させます。すると相互作用が始まってダイニンの運動が観察されます(図4)。のこぎり波様の信号がダイニン1分子の運動を示しています。詳細図では、ダイニンが階段を登るように動く様子を示しています。罫線は微小管の持つ周期構造に対応する距離8ナノメートルです。ダイニンへの負荷が大きいところでは、後ろ向きにずれる運動が頻繁に見られます。負荷が大きいほどこのずれの頻度は高くなっており、ダイニンが負荷に応じて運動を変化させていることがわかります。 これは力発生の調節を1つの分子が行うことを示したもので、タンパク質の自己調節機能を直接計測することに成功した世界で初めてのケースです。また、このような自己調節機構は鞭毛の波打ち運動においてダイニン間の協調を作り出すのに役立っていると考えられます。 | ||||
| ||||
| まとめ タンパク質モータは約2000~5000個のアミノ酸からなり、機能に関わる総原子数は10万~100万の範囲です。これは量子力学で扱うにはあまりにも大きな多体です。一方、タンパク質はモジュール構造と呼ぶ機能構造を持つために、統計力学の通常の扱いが難しい対象でもあります。 タンパク質機能を分子レベルで扱う領域は学問的に未開拓の領域です。近年の単一分子レベルでのタンパク質研究の進展、遺伝子改変によるタンパク質発現系の確立、X線結晶解析の技術革新によるタンパク質構造知見の拡大から、タンパク質機能の解明は大きく進むものと期待されています。ナノメートルの機能素子としてのタンパク質研究は新たな時代を迎えつつあり、私たちもその一端を担っています。(本成果はNatureに掲載されました。) (関西支所 生体物性研究室長) | ||||
 | ||||
| ||||
 液晶レンズの研究開発 液晶レンズの研究開発-宇宙通信機の小型化をめざして ― デジタル・ナロー通信方式とは? -400MHz帯等業務用無線の周波数有効利用の促進に向けて ― BOOK INFORMATION |