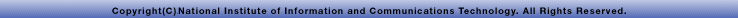背景
バイオICT研究室では生命が持つ巧妙な情報伝達の仕組みを解明し、未来の情報通信技術に役立てるための基礎研究に取り組んでいます。生命活動のすべてにおいて、環境の変化に応答する必要があり、それはDNAレベルにまで及びます。生物が有性生殖を選ばざるを得なくなったのは環境変化や外敵(病原菌など)との戦いで生き延びるための戦略と言われています。有性生殖の基本を司るのが減数分裂(ヒトでは卵子や精子をつくる過程)です。減数分裂においては、性の異なる個体の間で染色体DNAの交換が行われ、その結果、生物が遺伝子の多様性を獲得し、種の繁栄と継承を守ってきました。
私たちの細胞には父母それぞれから貰った2本(一対)の相同な染色体がありますが、減数分裂においては、この相同染色体の間で遺伝子交換(相同組換え)が行われます。バイオICT研究室の生物情報グループでは、染色体の挙動を生きた細胞で可視化する研究を行い、染色体の末端(テロメア)が近接したテロメアブーケ*1と呼ばれる特殊な構造が形成されること、それに引き続いて核運動がおこることが、相同染色体が空間的に近づくことに必要であることを明らかにしてきました。しかし、この過程で最も謎とされてきた部分は、相同染色体同士が、たくさん(ヒトの場合は全部で46本)ある他の染色体の中から、どのような仕組みで自分と同じ配列をもつ相同染色体だけを見分けて横に並ぶか(対合するか)は謎のままでした。今回の研究では、減数分裂を簡単に誘導できる分裂酵母を使ってこの謎を解く糸口を見つけました。
高頻度に対合する染色体サイト
分裂酵母では、Mei2*2という減数分裂を制御するタンパク質が存在し、そのタンパク質は減数分裂期前期に、染色体の決まった場所(sme2*2遺伝子座)に集積体を作ります。細胞には相同なsme2遺伝子座が2つあるにもかかわらず、観察されたMei2集積体が1つしかないことから、sme2遺伝子座が互いに対合しやすい可能性が示唆されました。sme2遺伝子座の対合を調べた結果、これまでに調べたほとんどの染色体領域よりも高い対合率を示すことが分かりました(図1)。さらに、sme2遺伝子を別の染色体に移すと、元の場所での対合頻度が減少し、移された場所の対合頻度が上昇したことから、sme2遺伝子座が相同染色体の対合を強く促進することが分かりました。しかも、sme2遺伝子座の対合はテロメアブーケの形成と核運動に依存することも分かりました。これらの結果は、まず、テロメアブーケの形成と核運動によって相同染色体同士が近接した位置に配置され、次にsme2遺伝子座の働きによって相同染色体が対合することを意味します。
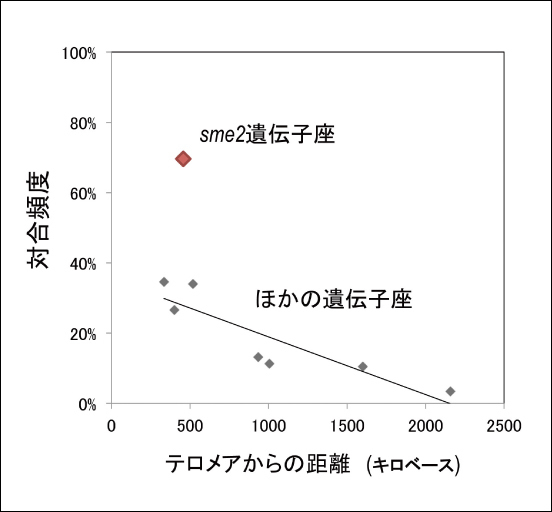
図1●高い対合頻度を示すsme2遺伝子座減数分裂期前期のsme2遺伝子座の対合頻度はほかの遺伝子座よりはるかに高い。
相同染色体双方のRNA転写は対合の鍵
それでは、なぜsme2遺伝子座には染色体の対合を促進する働きがあるのでしょうか。sme2遺伝子はタンパク質の設計情報を持たない非コードRNAを産生します。このRNA 産生を阻害すると対合頻度が減少することから、sme2から作られる RNA産物が対合促進に必須であることが分かります。さらに、相同染色体の片方だけからの転写では対合促進が起こらないことから、相同染色体の双方からのRNA 産生が対合に必要であることが分かりました。このRNAの局在を可視化すると、染色体上のsme2遺伝子座に集積していることが分かりました(図2)。また、sme2遺伝子の末端を欠損すると、RNAがsme2遺伝子座に留まることができなくなり、対合も阻害されることが分かりました。これらの解析から、我々は、sme2遺伝子座に滞留する RNAの集積が、相同染色体を見分けて優先的に対合を引き起こすメカニズムの実体であることを明らかにしました。
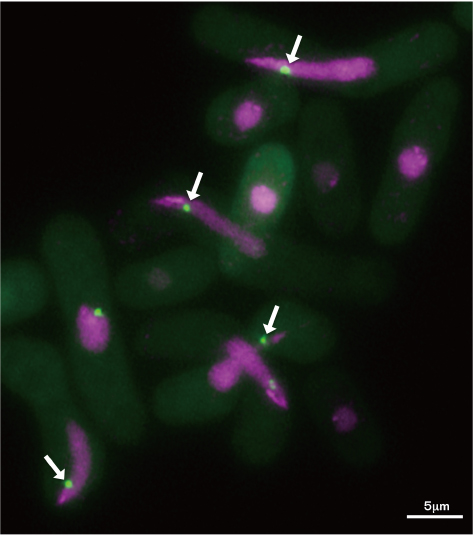
図2●sme2遺伝子座に局在する非コードRNA分裂酵母減数分裂期前期核(DNA: マジェンダ)にあるsme2遺伝子座に滞留する非コードRNA集積体(緑)(矢印)の生細胞観察写真。
今後の展望
今回の染色体上に蓄積する非コードRNAが染色体の認識に寄与するという新しい発見は、相同染色体認識のメカニズムを世界で初めて明らかにしたものです。対合するべき相同染色体を識別するために、損傷が致命的なエラーにつながるDNAそのものではなく、DNAを鋳型にして多コピー合成できるRNAを利用することは、非常に合理的であると考えられます。また、染色体上の幾つかの場所でこのような非コードRNAの集積があれば、バーコードのように識別しやすい特徴を染色体に与え(図3)、相同染色体の認識を効率よく実現できる染色体の“お見合い”戦略と考えられます。今後は、非コードRNAが相同染色体対合を促進する分子メカニズムを解明すると共に、RNA等の生体分子を利用したバイオセンサー等の情報通信技術への応用につなげていきます。
図3●相同染色体対合のモデル図テロメアブーケの形成と共に、染色体数ヶ所からRNAを含む対合サイトが形成され、染色体にバーコードのような識別特徴を与え、相同染色体の相互認識と対合を促進する。
なお、この成果は国際的科学誌「Science」2012年5月11日号に掲載されました。
用語解説
*1 テロメアブーケ
減数分裂期前期にすべての染色体末端テロメアが、核膜の1ヶ所に集まり、染色体を花束のように束ねる現象で、酵母から哺乳類、植物まで共通に見られる現象です。テロメアブーケの形成およびテロメアを先頭とする核運動によって相同染色体が同じ方向に並べられ、空間的に近づけられるようになり、対合が効率よくできるようになります。
*2 Mei2,sme2
Mei2は減数分裂を制御する遺伝子の名前です。sme2はsuppressor of mei2の略。sme2遺伝子からはmeiRNA-Lという非コードRNAが転写され、それがsme2遺伝子座上に蓄積され、相同染色体の対合を強く促進する働きがあることが我々の研究で明らかになりました。
 |
丁 大橋(テイ ダイキョウ) 未来ICT研究所 バイオICT研究室 主任研究員 大学院修了後、1992年、郵政省通信総合研究所(現NICT)に入所。生物情報の可視化技術の開発、相同染色体の認識・対合機構の研究に従事。博士(理学)。 |
![]() 1208号_3-4p(印刷用、570KB、A4 2ページ)
1208号_3-4p(印刷用、570KB、A4 2ページ)